
Extract from pages 6 - 23 in:
GRASSES of New South Wales
by
D.J.B. WHEELER (formerly Botany, University of New England)
S.M.L. JACOBS, National Herbarium, Royal Botanic Gardens, Sydney. N.S.W
R.D.B. WHALLEY Botany, School of Environmental Sciences and Natural Resources Management, University of New England.
THE UNIVERSITY OF NEW ENGLAND ARMIDALE AUSTRALIA
Reproduced at www.fog.org.au with written permission from The University of New England
©The University of New England, 1982
This work is copyright. Apart from any fair dealings for the purpose of private study, research, criticism, or review, as permitted under the Copyright Act, no part may be reproduced by any process without written permission. Inquiries should be made to the publishers.
Published by Botany School of Environmental Sciences and Natural Resources Management University of New England Armidale, NSW, 2351 Australia
Reprinted 1984 2nd edition 1990 3rd edition 2002
THE GRASS PLANT
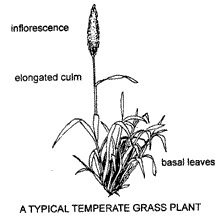
Grasses commonly grow in tufts or tussocks, each plant consisting of a cluster of leafy shoots, each shoot being termed a tiller. In a tuft the tillers are joined at the base by very short stems and this growth habit is known as caespitose or tufted. However, tillers can be arranged in other ways. In mat grasses, shoots are attached to long, often branched, stems growing alone the ground as illustrated. Other growth habits are described on page 12.
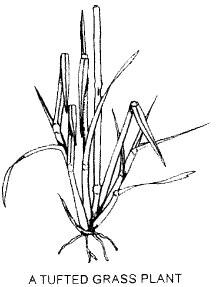

Tillers
A grass tiller consists of roots, stem and leaves, the most obvious part being the leaves. These are elongated structures made up of a basal cylindrical sheath and an upper blade (lamina) that may be flat, folded or rolled with overlapping edges. Each leaf is attached to a specialised portion of the stem, a node, where the veins of the leaf are integrated with the vascular system of the stem. Between the nodes are internodes as shown in the illustration.
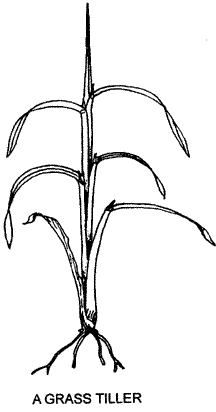

A longitudinal section (LS.) of the basal portion of a tiller of a temperate grass plant, such as Phalaris aquatica (illustrated below) shows the arrangement of leaves in the vegetative stage. The cylindrical leaf-sheaths are attached to the stem nodes with very short internodes in between. The older leaves encircle and protect the younger leaves and the apical bud. By being very close to the ground this bud is also protected from grazing animals. An axillary bud may be seen in some lower leaf axils (the axil is immediately above where the leaf sheath joins the node). Such buds can develop into new tillers, as has the bud in a leaf axil on the right of the diagram. Roots arise from the lower nodes of grass tillers.
Shoots with very short internodes at or near ground level, with leaves rising high above them and acting as a protective sheath, are characteristic of the vegetative stage of most grasses. It is usually only when the tiller is ready to flower that the internodes elongate carrying the apical bud above the leaves to produce an inflorescence or seedhead. Each tiller usually dies after flowering and fruiting. In annual grasses the whole plant dies, while in perennials the growth of the plant over subsequent seasons is continued by the formation of new tillers. The individual tillers may live from one to several years.

Leaves
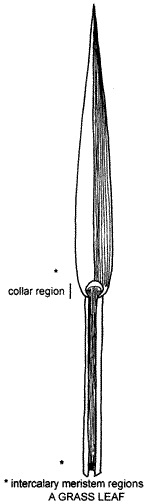 The
leaf of a grass is an elongated structure arising at a node and consisting of a
basal cylindrical sheath that encircles the stem or younger leaves. Above this
is a flattened blade or lamina with parallel venation. The parallel veins are
drawn on the right hand side of the blade. The leaves are borne alternately on
either side of the stems. One exception is the tropical Australian genus
Micraira with spirally arranged leaves.
The
leaf of a grass is an elongated structure arising at a node and consisting of a
basal cylindrical sheath that encircles the stem or younger leaves. Above this
is a flattened blade or lamina with parallel venation. The parallel veins are
drawn on the right hand side of the blade. The leaves are borne alternately on
either side of the stems. One exception is the tropical Australian genus
Micraira with spirally arranged leaves.
Leaf-sheaths are hollow cylinders split down one side with the margins usually overlapping. In some grasses the margins may be joined together (fused) to form a complete cylinder as in Bromus spp.
Leaf-sheath features useful in identification are the degree of flattening or compression, hairiness, and sometimes the colour and prominence of the veins or nerves. At the base of the leaf sheath and the blade are intercalary meristems that, for all but old leaves, allow the leaf to elongate after removal of the blade by mowing or grazing or damage by wind and rain. Intercalary meristems are regions in which cells retain their ability to divide and produce new cells. Because grass leaves mature from the tip of the blade down to the base, the sheath meristem collar region remains active longer than the blade meristem. In grasses, intercalary meristems also occur at the base of stem internodes where activity enables the stem to grow upright after lodging.
Though the shape, texture, folding, hairiness etc. of the leaf-blade may be useful diagnostic features, they are often variable within a species or even on the same plant. In some leaf-blades the midrib is prominent, and often pale-coloured, while in others the veins are equally conspicuous or inconspicuous. Leaf-blades are often flat but may be rolled or folded, (rolling or folding is a response to water stress in some cases) and some are so narrow as to be bristle-like. A band of mechanical tissue, often pale, occurs at the blade/sheath junction, the abaxial portion of which is termed the collar. (Abaxial refers to the surface intercalary meristem regions facing away from the axis of the plant). An abscission layer may also develop enabling the blade to be shed.

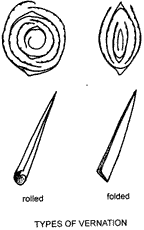
Vernation, the arrangement of a young leaf in the bud, is a very useful diagnostic feature, often remaining constant for a genus e.g. Austrodanthonia. Young leaves may be either folded, as in Austrodanthonia where the edges lie side by side, or rolled with the edges overlapping, e.g. Oats, Avena sativa. Transverse sections (TS.) of tillers and views of leaves from the outside are illustrated.
A ligule, in the form of hairs or a tongue or flap-like transparent membrane, is attached on the leaf surface towards the axis (adaxial) at the blade/sheath junction (or collar). Ligule type is a particularly useful identification character as it is relatively constant within a genus. Some grasses may lack ligules e.g. Echinochloa crusgalli, Barnyard Grass. Many grasses have translucent, membranous ligules whereas others have ligules fringed with hairs or in the form of a rim of hairs.

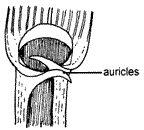 Ear-like
projections, auricles, are often produced at the top of the leaf-sheath.
Auricles may be reduced to a hairy edge of the leaf-blade base or may occur on
one side of the leaf only, in which case it is termed an auricular lobe.
Auricles, though a distinctive feature of many species, are delicate structures
and may wither with age. Young leaves should always be examined to determine
auricle shape.
Ear-like
projections, auricles, are often produced at the top of the leaf-sheath.
Auricles may be reduced to a hairy edge of the leaf-blade base or may occur on
one side of the leaf only, in which case it is termed an auricular lobe.
Auricles, though a distinctive feature of many species, are delicate structures
and may wither with age. Young leaves should always be examined to determine
auricle shape.

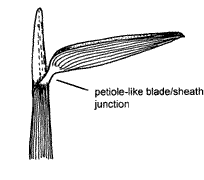 In
most bamboos and some tropical grasses e.g. Setaria palmifolia, there is a
petiole-like constriction of the blade/sheath junction. The length of the
constricted area varies from species to species (e.g. Stenotaphrum secundatum, Buffalo Grass, in which the constriction is very short).
In
most bamboos and some tropical grasses e.g. Setaria palmifolia, there is a
petiole-like constriction of the blade/sheath junction. The length of the
constricted area varies from species to species (e.g. Stenotaphrum secundatum, Buffalo Grass, in which the constriction is very short).
Modified leaves
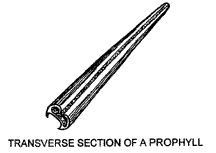
A new shoot, if produced within the subtending leafsheath by development of an axillary bud, is termed intravaginal and is protected by a prophyll. The prophyll (in grasses) is a two-keeled, modified leaf without a blade, attached to the node on the side towards the parent axis. Prophylls subtend side shoots where branching occurs at aerial nodes or on many horizontal stems.
The diagrams show a prophyll in section and its position on the plant.
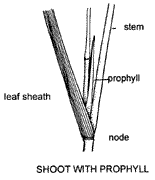
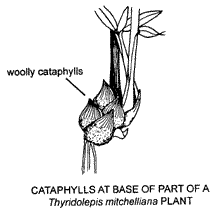
New shoots often break through the subtending leafsheath in which case they are termed extravaginal. The first formed leaves of tillers of many tufted grasses e.g. Mulga Grass (Thyridolepis mitchelliana), are modified and termed cataphylls. Presence of cataphylls is often an indication of extravaginal branching. Cataphylls usually closely overlap each other, do not have blades, and may be a distinctive feature. They are succeeded distally by normal leaves (i.e. normal leaves are produced along the stem above the cataphylls).
Stems
The upright stems of grass tillers are termed culms. They are solid at the nodes and often hollow throughout the internode region. However, the internodes of many grasses, such as Sugarcane, are pithy while those of many aquatic grasses, such as Rice, have a series of inter-connected air chambers in addition to the central cavity.
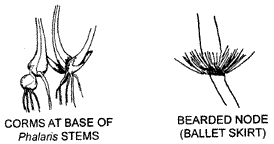
Storage of carbohydrate frequently occurs in grass culms, for instance in Sugarcane. Storage also takes place in the basal portion of many grass stems. In such species as Panicum bulbosum and Phalaris aquatica the basal internodes become enlarged to form corms as illustrated.
Nodes may he useful distinguishing features. They may be relatively inconspicuous, enlarged, pigmented or fringed with hairs. The 'ballet skirt' of Sorghum leiocladum is illustrated below.
Culms of most temperate grasses do not elongate until just before flowering whereas those of some tropical grasses such as Saccharum officinarum (Sugarcane), are elongated in the vegetative stage. Many grasses are somewhat intermediate in habit, having much-branched stems with clusters of leaves and buds at various heights above branched the ground, in contrast to the typically basal leaves and culms buds of vegetative temperate grasses (see illustrations on this page and p. 12). Grass buds on the much-branched stems are not as well protected from grazing animals as are the basal buds of temperate grasses. Animals also take longer to obtain leafy fodder from tall, diffuse tropical grass stands than from temperate pastures with basal concentrations of leaves. The three plants illustrated (two on this page and one on p. 12) are examples of tufted perennials. Such plants may form large tussocks if allowed to grow for many years. Growth in plant diameter is a result of the production of new tillers from basal buds.

 Growth
habits
Growth
habits
If the lower nodes of a grass culm lie near the ground, roots may be produced at these nodes. Such a plant is often described as decumbent Many grasses produce horizontal stems, usually by extravaginal branching, that grow above or below the ground. Such stems are termed stolons if they grow above the ground and rhizomes if growth occurs below ground. Stoloniferous grasses produce green leaves and tillers at each node, while brown, reduced scale leaves are attached to the nodes of rhizomes. Both stolons and rhizomes root at the nodes and, if the internodes between the rooted nodes are broken, each piece can develop into a new plant. Rhizomatous and stoloniferous grasses are able to reproduce very efficiently in the vegetative stage and can spread rapidly over or through the ground.
Most stoloniferous and many rhizomatous grasses are termed mat grasses. Some such as Couch (Cynodon dactylon) are useful for lawns and pasture. The ability of grasses such as Couch and Kikuyu grass (Pennisetum clandestinum) to produce both stolons and rhizomes also makes them troublesome garden weeds. Some grasses produce long trailing stems that climb long distances over other vegetation, an example being Tetrarrhena juncea.
Though most grasses are herbaceous some are shrubby with the culm bases becoming woody. Bamboos have perennial woody stems that may reach 30 m tall. Some grasses of the Australian inland form hummocks a metre or more in diameter. As the diagram shows, branched stems occur in the centre of the hummock with green leaves at the tips. The leaf tips of certain hummock grasses such as some Spinifex (Porcupine) grasses (Triodia spp.) are needle sharp. Hummocks are also sometimes described as tussocks.


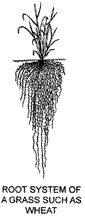 Roots
Roots
Grass roots are much branched, do not become enlarged for storage and support, and are termed fibrous or adventitious. The root system of most grasses is extensive, the ratio of roots to tops by weight varying from 0.7: 1 to 4: 1. High proportions of roots are characteristic of plants growing in drier habitats.
The first-formed roots of a grass seedling arise from the embryo and form the primary or seminal root system. Such roots are important in seedling establishment. Later, roots are produced from the lower nodes of the young plant and its tillers. These roots become increasingly important, finally forming the mature root system. The soil-binding properties of grasses are related to the extensive network of fine roots.
Inflorescence
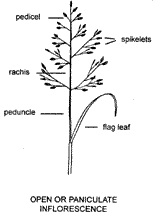 The
stem of a grass tiller in the reproductive stage terminates in an inflorescence
or flowering head. Each inflorescence consists of a number of spikelets, which
contain the very small flowers, arranged on branches or the main axis.
Spikelets, the units of the grass inflorescence, are either attached directly to
the branches, in which case they are sessile, or are attached by means of stalks
or pedicels. The stem immediately below the inflorescence is the peduncle. which
continues as the main axis or rachis after the first branch or spikelet.
'Rachis' (sometimes as 'rhachis') is a general term and often used for any axis
or branch to which spikelets are attached. The uppermost leaf, often partially
enclosing the inflorescence, is termed the flag leaf if it is not particularly
differentiated from the other leaves. If it is differentiated from the other
leaves, and subtends an inflorescence, it is called a spathe.
The
stem of a grass tiller in the reproductive stage terminates in an inflorescence
or flowering head. Each inflorescence consists of a number of spikelets, which
contain the very small flowers, arranged on branches or the main axis.
Spikelets, the units of the grass inflorescence, are either attached directly to
the branches, in which case they are sessile, or are attached by means of stalks
or pedicels. The stem immediately below the inflorescence is the peduncle. which
continues as the main axis or rachis after the first branch or spikelet.
'Rachis' (sometimes as 'rhachis') is a general term and often used for any axis
or branch to which spikelets are attached. The uppermost leaf, often partially
enclosing the inflorescence, is termed the flag leaf if it is not particularly
differentiated from the other leaves. If it is differentiated from the other
leaves, and subtends an inflorescence, it is called a spathe.
The basic type of grass inflorescence is a compound panicle, usually referred to simply as a panicle, with secondary and often higher order branching. A spike is an inflorescence with a single axis and spikelets attached directly, while in a raceme the spikelets are attached to the rachis by pedicels. Many inflorescences are intermediate in type and may be thought of as panicles with reduced branching. If the branches are short and crowded together, the panicle is narrow and described as spike-like, spicate, contracted or dense. Reduction in the number of branches can give rise to a primary axis with racemes or spikes. If the axis is very short, such a panicle is called sub-digitate. Also included is the digitate type in which racemes or spikes are arranged on the top of the peduncle like fingers of a hand.
An approach, used in many of the descriptions in this book, is to describe inflorescences in categories that are increasingly inclusive. For instance, the terms 'spike', 'spicate' and 'spike-like panicle' are increasingly more inclusive. Spicate can include all spikes but spike cannot include all spicate inflorescences. Similarly, 'spike-like panicle' can include all of the other two. The term 'racemose' can be used in a similar way with racdemose including racemes and raceme-like inflorescences but raceme being restricted to those that are strictly racemes. 'Paniculate' can be used as a general term for any branched inflorescence.
Leaves are not usually found in grass inflorescences but, in some tropical grasses, the inflorescence is a complex collection of branches with a leaf-like structure, a spathe, at the base of each branch or section of the inflorescence. The branches may be condensed or modified and are often in pairs, as is the case in the illustration where racemes are in pairs above the spathes. The spathes may be coloured or green and may or may not be differentiated into blade and sheath.
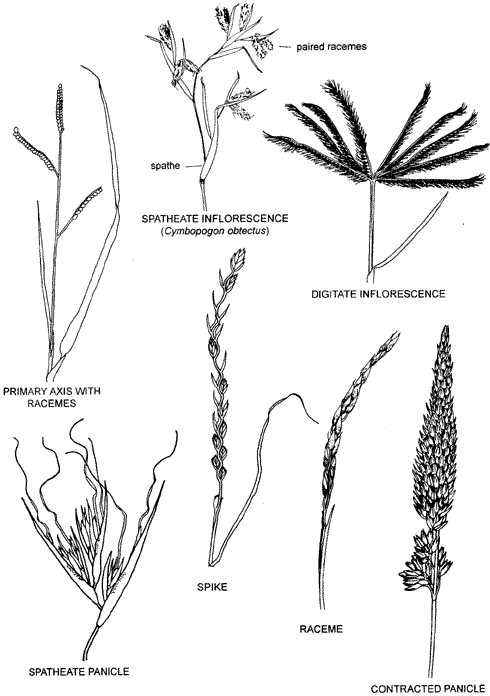

Examples of some grass inflorescences: Primary axis with racemes (Paspalum); digitate inflorescence (Chloris); spike (Lolium); spatheate panicle (Themeda); raceme (Zoysia); contracted panicle (Phalaris); open panicle (Panicum).
Spikelets
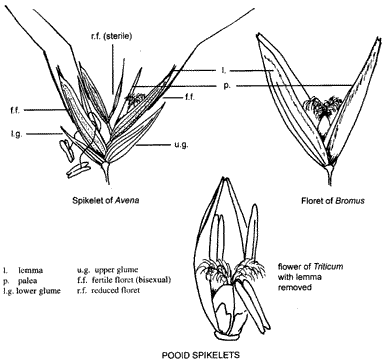
Each spikelet consists of a number of flowers or florets arranged alternately on either side of a central axis, the rachilla, (sometimes as 'rhachilla') above two empty bracts, the glumes. 'Bract' is a general term for a much reduced leaf, particularly small leaves subtending flowers or inflorescences. A grass flower is enclosed by two bracts, the outer one on the side away from the axis termed the lemma and an inner bract named the palea. The bracts, together with the flower, make up the floret. The palea, because it is the bract just below the flower is sometimes termed a bracteole (= small bract) (see also p. 16).

Glumes
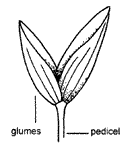 Each
spikelet has two glumes at the base. The glumes never subtend a flower or palea.
They may, however, be modified and vary in size, shape, texture, etc. and one
(or rarely both) may be absent. Looking like stripes on the glumes, are strands
of vascular tissue (veins) termed nerves. The number and prominence of nerves is
variable between species, but reasonably constant within species. Glumes may
resemble the lemmas above them and it is important, when examining grass
inflorescences, to be able to recognise the glumes and thus identify spikelets
as units.
Each
spikelet has two glumes at the base. The glumes never subtend a flower or palea.
They may, however, be modified and vary in size, shape, texture, etc. and one
(or rarely both) may be absent. Looking like stripes on the glumes, are strands
of vascular tissue (veins) termed nerves. The number and prominence of nerves is
variable between species, but reasonably constant within species. Glumes may
resemble the lemmas above them and it is important, when examining grass
inflorescences, to be able to recognise the glumes and thus identify spikelets
as units.
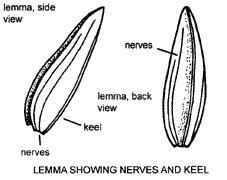
Florets
The outermost structure of the floret is the lemma, which usually encloses the other parts. If laterally compressed, a lemma typically has a single ridge or keel running down the middle of the back. Usually visible, and often constant in number for a particular genus or species, are several veins or nerves. Lemmas are frequently green and membranous, especially when immature, and become harder and often less green as they mature. There is considerable variation in size, shape, colour, texture, compression, hairiness and presence of appendages such as awns (see p. 18). Such variations are particularly useful in identification.
At the base of many florets is a hardened structure, often sharply pointed, that develops at maturity and can be seen clearly when the floret separates from the rachilla.
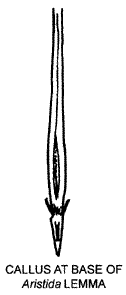
'Callus' is the term used for the proximal end of the diaspore (sexual dispersal unit) in grasses, the end closest to its subtending axis. Ideally its use is restricted to those grasses where the diaspore is either a floret or a spikelet, the situation for the vast majority of species. In some grasses the caryopsis behaves as the diaspore (e.g., Eragrostis, Sporobolus), in others it is a group of spikelets (e.g., Phalaris paradoxa, Cenchrus, Sorghum), and there are others where the inflorescence breaks into segments with embedded spikelets (e.g., Hemarthria, Hainardia, Stenotaphrum); it is probably best not to use 'callus' in these cases. In those species where the floret disarticulates above the glumes and between the florets (e.g., most of the Pooid and Chloridoid grasses), the callus consists of lemma and rachilla tissue, with possibly some palea tissue. In those species where the disarticulation is below the glumes (e.g. most Panicoid grasses) the callus consists of glume and pedicel tissue. In some Andropogonoid grasses the spikelets may fall in pairs or triplets (e.g. some species of Sorghum, Chrysopogon) and, if the rachis part is pungent, sometimes it may be (though not ideally) referred to as a callus.
Because the term may be used for essentially different tissues, it is basically a term defined on perceived function and not on homology and care is needed when this character is used in any analyses. The callus may be hard, sharp and penetrating (e.g., Austrostipa spp., Oxychloris, Heteropogon), blunt and obtuse (e.g., Deyeuxia, Poa, Sacciolepis) or anything in between (e.g., Austrodanthonia, Bothriochloa, Dichelachne). Where the callus is hard, sharp and penetrating it frequently has stiff antrorse hairs, and often the diaspore also has a geniculate awn. This syndrome is characteristic of species that bury their diaspores in the ground. These same species may also penetrate skin and socks, and catch on fur or clothing. Contrary to popular belief., these are not adaptations for spread as the diaspore is usually never released from its 'target'. The syndrome appears to have developed well before fur and clothes were common.
Paleas are usually 2-keeled (and 2-nerved) and softer and less variable in structure than lemmas. In the floret, a flat side lies towards the rachilla while the keels touch the sides of the lemma, thus enclosing the flower. Paleas of bamboos and Rice, Oryza sativa, and their relatives are only partly enclosed in their lemmas.
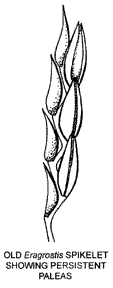

Rachilla
This small axis is bent alternately in opposite directions (flexuose or "zig-zag") and bears the florets, which are in two rows. This is shown in the diagram of part of an old Eragrostis spikelet where all other floret parts have fallen, leaving the paleas with their backs to the zig-zag rachilla.
Flowers
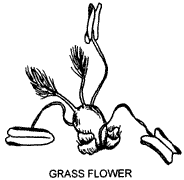

As can be seen from the diagram, there are no sepals or petals but 2-3 small fleshy structures, the lodicules, at the base of the ovary. These are considered to represent a reduced perianth. The lodicules are delicate structures lying between the ovary and the lemma (and palea, if a third is present). As the flower opens (anthesis) they swell, pushing the lemma and palea apart to allow first the anthers (usually) and then the stigmas to be exserted (project beyond the bracts) allowing pollination. Protandry is the condition in which the stamens are exserted first and protogyny when stigmas appear before anthers. When stigmas have been exposed for several hours, the lodicules shrivel, allowing lemma and palea to close over the developing grain. Two types of lodicules are shown in the diagram and the names refer to two major groups of grasses. Pooid grasses grow mostly in temperate regions, an example being Wheat, Triticum aestivum, while panicoid grasses are usually tropical, for instance Maize, Zea mays, or Paspalum, Paspalum dilatatum.
Attached at the base of the flower, in a ring or whorl inside the lodicules, are 1-6 stamens. Most grasses have 3 stamens, as illustrated, with long delicate filaments. The anthers, which open by means of slits, are versatile, that is the filaments are attached at one point above the base. This allows the anther to swing with the wind enabling pollen to be shed in all directions.
The usually elongated superior ovary contains a single loculus or chamber, with one ovule attached near the base on the palea side. At the apex of the ovary are two lateral styles (in some grasses fused to give a single central style) bearing hairy stigmas. In pooid grasses the stigmas are pale while those of panicoid grasses are usually dark (pink to black).
Fruits
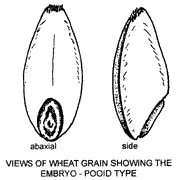
The ovule of a grass flower develops, after fertilisation, into a seed with much starchy endosperm and an embryo lying towards one side. The ovary wall, which forms the pericarp of the fruit, is normally fused with the seed coat and the fruit termed a caryopsis or grain. Sporobolus and Eleusine produce achenes in which the testa (seed coat) is free from the pericarp. Some bamboos produce berries. Most grass grains are shed enclosed in the lemma and palea. Shape, surface texture, relative size and position of the embryo are useful diagnostic features.

Variations in spikelet structure
Many grass taxonomists consider the basic spikelet pattern to be similar to the many-floreted spikelets of grasses such as Oats, Avena sativa, illustrated on p. 20. This is thought to have been modified by addition of such structures as awns and hairs etc. and by reduction of the numbers and/or size of spikelet parts. The basic spikelet pattern is also the one shown in the diagram on p. 15.
Additions
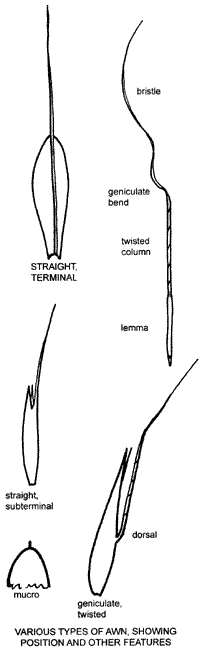 Hairs,
bristles, spines etc. may be attached to parts of spikelets. In some cases such
structures surround individual spikelets or groups of spikelets (see p. 23).
However, the most obvious appendage is the awn.
Hairs,
bristles, spines etc. may be attached to parts of spikelets. In some cases such
structures surround individual spikelets or groups of spikelets (see p. 23).
However, the most obvious appendage is the awn.
An awn is a long slender appendage attached to the tip, back or base of a glume, lemma or palea, the point of attachment being a diagnostic feature. Some awns are spirally twisted, many twisting and untwisting in response to changes in humidity thus enabling the base of the lemma (often with a sharp callus) to penetrate soil or hides. Such awns are divided into two parts, a relatively stout column and a terminal, slender bristle. A very slender awn is often termed a bristle and a very short, terminal awn, a mucro.
Reductions
 Both
the number and the size of spikelet parts can be reduced. The number of florets
in a spikelet varies from 1-20 and, in some grasses, spikelets are reduced to
glume(s) only.
Both
the number and the size of spikelet parts can be reduced. The number of florets
in a spikelet varies from 1-20 and, in some grasses, spikelets are reduced to
glume(s) only.
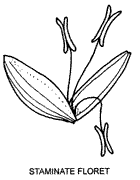
If the ovary alone is missing, the floret is termed staminate or male although it is often also called infertile or sterile. A floret without stamens or ovary, with lemma and palea or with lemma only, is termed sterile or neuter, although the terms infertile or empty are also used. A floret with an ovary is described as fertile. The terms sterile, fertile, etc. may be applied to lemmas of appropriate florets.
Although grass florets are normally bisexual, unisexual flowers, both male and female, are not uncommon. They may occur in spikelets with bisexual florets, in single-sex spikelets or in inflorescences with spikelets of one sex only. In monoecious grasses male and female inflorescences occur on the same plant, e.g. Maize, while male and female inflorescences occur on separate plants in dioecious grasses e.g. Spinifex sericeus (see p. 22).

The size of glumes, lemmas and paleas may also be reduced. In Weeping Grass, Microlaena stipoides, the glumes are reduced to tiny scales and in Ryegrass, Lolium spp., one glume is absent on all but the terminal spikelet. The sterile lemmas of some Phalaris spp. may be 1 mm in length.
Other variations
The way in which spikelets break up at maturity can be a useful diagnostic feature. A joint (abscission layer) may be present on the rachilla above the glumes (as shown by the upper arrow below) allowing the florets to fall at maturity leaving the glumes on the pedicel. Alternatively the glumes may fall with the spikelet, in which case the spikelet is said to disarticulate below the glumes. Disarticulation may also take place at joints of the rachilla and of the rachis. The paired spikelets of grasses belonging to the tribe Andropogoneae usually fall as a unit together with a section of the rachis of the raceme to which they are attached.
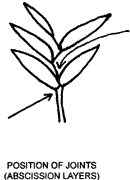

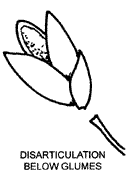

The rachilla is continued beyond the uppermost floret as a small bristle in some pooid spikelets. This 'prolongation' of the rachilla, often difficult to see, is useful in identification.
The position of reduced florets in a spikelet is also important. Reduced florets may occur on the rachilla above or below fertile (perfect) florets.
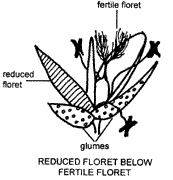

Several of the variations mentioned tend to occur together and characterise two major groups of grasses. Pooid grasses, with the basic spikelet pattern shown in the diagram on p. 15, have 1-many florets per spikelet with reduced florets above fertile florets and disarticulation above the glumes. Panicoid grasses exhibit what is often termed a 'reduced' spikelet pattern. In these grasses the floret number is reduced to 2, with a sterile (reduced) floret below a fertile floret on a very short rachilla, and disarticulation is below the glumes. There are of course exceptions to these generalisations. Differences in spikelet morphology, though still most useful in grass identification, have declined in importance for classification as information on anatomical, physiological and other such characters has become available in recent years.

The degree of maturity of spikelets is likely to vary within a large inflorescence. Often the older spikelets are towards the top or middle of the inflorescence. It is desirable, and usually possible, to examine spikelets at different stages of maturity when identifying a grass.
Spikelet arrangement
Spikelets in an inflorescence are usually similar in appearance but 2 types may occur in the same inflorescence or in different inflorescences. In Lamarckia (illustrated) sterile spikelets, not producing fruits, are mixed with fertile spikelets.

Spikelets of many genera of the tribe Andropogoneae are borne in dissimilar pairs, one member of the pair being fertile (and often sessile) and the other pedicellate, sterile and smaller.
Male and female spikelets may occur in separate inflorescences which are often unlike in appearance. In a monoecious plant like Zea mays, both types of inflorescence are found on the same plant. The male inflorescence is terminal and the female (the cob) is in the axils of the lower leaves.
In a dioecious plant such as Spinifex sericeus male and female inflorescences are on different plants. (See illustration). In Cleistochloa, spikelets with flowers that open normally for pollination (chasmogamous) are in one type of inflorescence, while cleistogamous spikelets (which do not open) are in inflorescences enclosed in the leaf-sheaths. Spikelets or inflorescences that occur in two forms are often termed dimorphous.



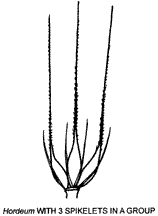
Many grasses, other than the andropogonoid types mentioned, have spikelets arranged in groups e.g. Hordeum (illustrated). Spikelets, or groups of spikelets, may be subtended by various structures that function in seed dispersal. These include hairs, bristles, spines, burrs or sterile branches. The bristle ring and burr of two Cenchrus species are illustrated.
The surface (epidermis) of leaves, stems, spikelets etc., may bear such structures as, hairs, tubercles, papillae and glands. These surface features also include patterns such as dots or fine lines. For more information see Glossary entries and particularly pp. 418 and 421. Glands are secretory structures that are distinguished by colour and texture from surrounding tissue. They are useful diagnostic features in genera such as Eragrostis.


Additional reading
Additional reading for the Grass Family and its Importance and The Grass Plant:
BARNARD, C. (Ed.) (1964) Grasses and grasslands.: (Macmillan: London)
CLIFFORD, H.T. & WATSON, 1. (1977). Identifying grasses, methods and illustrations.: (University of Queensland Press: St. Lucia)
COUPLAND, R.T. (Ed.) (1979) Grassland ecosystems of the world. I.B.P. 18. pp 8-35.( Cambridge University Press: Cambridge)
GOULD, F.W. (1968) Grass systematics. (McGraw Hill. New York)
HARLAN, J.R. (1975) Crops and man.: (American Society of Agronomy, Crop Science Society: Madison)
HEYWOOD, V11. (Ed.) (1978) Flowering plants of the world. [Gramineae pp 285-901: (Oxford University Press: Oxford)
JACKSON, D.C. & JACOBS, SW.L. (1985) Australian Agricultural Botany, 377 pp.: (Sydney University Press: Sydney)
MOORE, R. Milton (Ed.) (1970) Australian Grasslands: (Australian National University Press: Canberra)
PURSEGLOVE, LW. (1972) Tropical Crops: Monocotyledons (Longman: London)
SODERSTROM T.R., HILU, K.W., CAMPBELL, C.S. & BARKWORTH, M.E. (Eds.) (1987). Grass systematics and evolution. 474 pp. (Smithsonian Institution Press: Washington)